
Kladogram

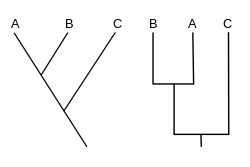
Kladogram (daripada bahasa Yunani clados "cabang" dan gramma "sifat") ialah gambar rajah yang digunakan dalam kladistik untuk menunjukkan hubungan antara organisma. Walau bagaimanapun, kladogram bukanlah pokok evolusi kerana ia tidak menunjukkan bagaimana nenek moyang terkait dengan keturunannya, dan juga tidak menunjukkan betapa banyaknya ia telah berubah, begitu banyak pokok evolusi yang berbeza boleh konsisten dengan kladogram yang sama.[1][2][3][4][5] Kladogram menggunakan garisan yang bercabang ke arah berbeza yang berakhir pada klad, sekumpulan organisma dengan nenek moyang terakhir yang sama. Terdapat banyak bentuk kladogram, tetapi semuanya mempunyai garisan yang bercabang dari garisan lain. Garis boleh dikesan kembali ke tempat ia bercabang. Titik bercabang ini mewakili nenek moyang hipotesis (bukan entiti sebenar) yang boleh disimpulkan untuk mempamerkan sifat yang dikongsi di antara takson terminal di atasnya.[4][6] Nenek moyang bayangan ini kemudiannya mungkin memberikan petunjuk tentang susunan evolusi pelbagai ciri, penyesuaian dan naratif evolusi lain tentang nenek moyang. Walaupun secara tradisinya kladogram sebegini dijana sebahagian besarnya berdasarkan aksara morfologi, data penjujukan DNA dan RNA dan filogenetik komputasional kini sangat biasa digunakan dalam penjanaan kladogram, sama ada secara sendiri atau bersama dengan penelitian morfologi.
Menghasilkan kladogram[sunting | sunting sumber]
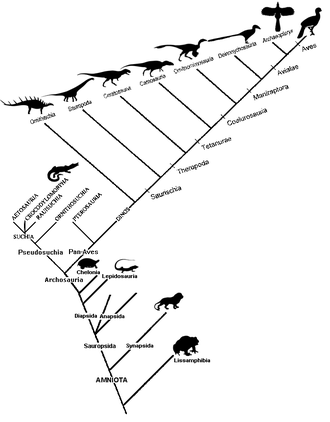
Data molekul lawan morfologi[sunting | sunting sumber]
Ciri-ciri yang digunakan untuk mencipta kladogram boleh dikategorikan secara kasar sebagai sama ada morfologi (tengkorak sinapsid, berdarah panas, notokord, unisel, dll.) atau molekul (DNA, RNA, atau maklumat genetik lain).[7] Sebelum kemunculan penjujukan DNA, analisis kladistik terutamanya menggunakan data morfologi. Data tingkah laku (untuk haiwan) juga boleh digunakan.[8]
Oleh kerana penjujukan DNA telah menjadi lebih murah dan mudah, sistematik molekul telah menjadi cara yang lebih dan lebih popular untuk membuat kesimpulan hipotesis filogenetik.[9] Menggunakan kriteria parsimoni hanyalah satu daripada beberapa kaedah untuk membuat kesimpulan filogeni daripada data molekul. Pendekatan seperti kemungkinan maksimum, yang menggabungkan model eksplisit evolusi jujukan, adalah cara bukan Hennig untuk menilai data jujukan. Kaedah lain yang berkuasa untuk membina semula filogeni ialah penggunaan penanda retrotransposon genomik yang dianggap kurang terdedah kepada masalah mutasi penerbalikan yang mengganggu data jujukan. Mereka juga secara amnya diandaikan mempunyai insiden homoplasi yang rendah kerana pernah difikirkan bahawa penyepaduan mereka ke dalam genom adalah rawak sepenuhnya; tetapi ini nampaknya sekurang-kurangnya tidak berlaku secara kadangkala.
Apa yang bukan kladogram[sunting | sunting sumber]
Kladogram ialah hasil rajah analisis, yang mengelompokkan taksa berdasarkan sinapomorf sahaja. Terdapat banyak algoritma filogenetik lain yang meneliti data dengan agak berbeza, dan menghasilkan pokok filogenetik yang kelihatan seperti kladogram, tetapi tidak. Sebagai contoh, algoritma fenetik seperti UPGMA dan penyambungan jiran, mengumpulkan berdasarkan persamaan keseluruhan, dan menganggap kedua-dua sinapomorf dan simplesiomorf sebagai bukti pengelompokan, dengan gambar rajah yang terhasil ialah fenogram, bukan kladogram, Begitu juga, hasil kaedah berasaskan model (kemungkinan maksimum atau pendekatan Bayes) yang mengambil kira kedua-dua tertib percabangan dan "panjang cawangan", mengira kedua-dua sinapomorf dan autapomorf sebagai bukti untuk atau menentang pengelompokan, dan gambar rajah yang terhasil daripada jenis analisis tersebut bukanlah kladogram juga.[10]
Lihat juga[sunting | sunting sumber]
Rujukan[sunting | sunting sumber]
- ^ Mayr, Ernst (2009). "Cladistic analysis or cladistic classification?". Journal of Zoological Systematics and Evolutionary Research. 12: 94–128. doi:10.1111/j.1439-0469.1974.tb00160.x.
- ^ Foote, Mike (Spring 1996). "On the Probability of Ancestors in the Fossil Record". Paleobiology. 22 (2): 141–51. doi:10.1017/S0094837300016146. JSTOR 2401114.
- ^ Dayrat, Benoît (Summer 2005). "Ancestor-Descendant Relationships and the Reconstruction of the Tree of Life". Paleobiology. 31 (3): 347–53. doi:10.1666/0094-8373(2005)031[0347:aratro]2.0.co;2. JSTOR 4096939.
- ^ a b Posada, David; Crandall, Keith A. (2001). "Intraspecific gene genealogies: Trees grafting into networks". Trends in Ecology & Evolution. 16 (1): 37–45. doi:10.1016/S0169-5347(00)02026-7. PMID 11146143.
- ^ Podani, János (2013). "Tree thinking, time and topology: Comments on the interpretation of tree diagrams in evolutionary/phylogenetic systematics" (PDF). Cladistics. 29 (3): 315–327. doi:10.1111/j.1096-0031.2012.00423.x. PMID 34818822 Check
|pmid=value (bantuan). - ^ Schuh, Randall T. (2000). Biological Systematics: Principles and Applications. ISBN 978-0-8014-3675-8.[halaman diperlukan]
- ^ DeSalle, Rob (2002). Techniques in Molecular Systematics and Evolution. Birkhauser. ISBN 978-3-7643-6257-7.[halaman diperlukan]
- ^ Wenzel, John W. (1992). "Behavioral homology and phylogeny". Annu. Rev. Ecol. Syst. 23: 361–381. doi:10.1146/annurev.es.23.110192.002045.
- ^ Hillis, David (1996). Molecular Systematics. Sinaur. ISBN 978-0-87893-282-5.[halaman diperlukan]
- ^ Brower, Andrew V.Z. (2016). "What is a cladogram and what is not?". Cladistics. 32 (5): 573–576. doi:10.1111/cla.12144. PMID 34740305 Check
|pmid=value (bantuan).
Pautan luar[sunting | sunting sumber]
 Kategori berkenaan Kladogram di Wikimedia Commons
Kategori berkenaan Kladogram di Wikimedia Commons