
Gliserofosfolipid

Gliserofosfolipid atau fosfogliserida ialah fosfolipid berasaskan gliserol. Ia merupakan komponen utama membran biologi dalam sel eukariot. Ia merupakan sejenis lipid, di mana komposisinya mempengaruhi struktur dan sifat membran.[1] Dua kelas utama diketahui: kelas bakteria dan eukariot, dan keluarga yang berasingan bagi arkea.[2]
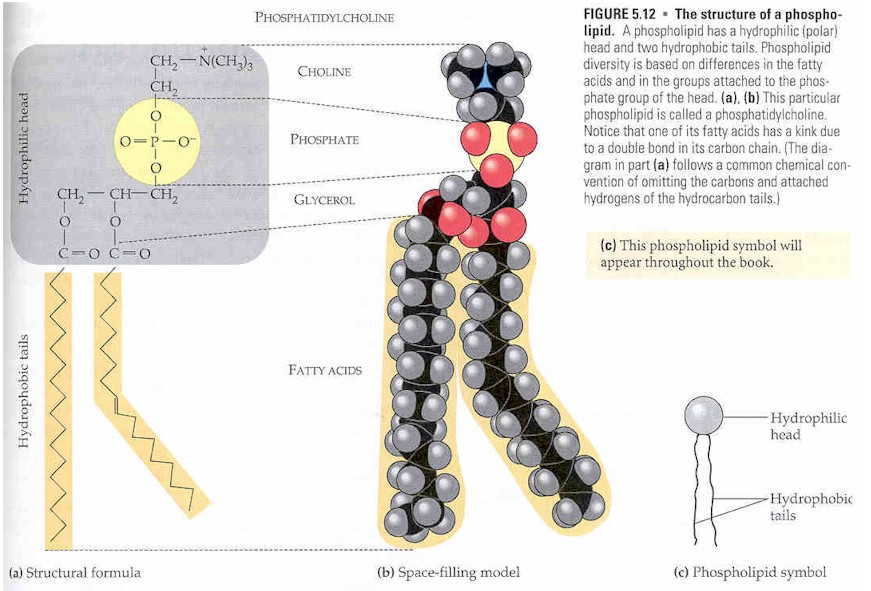
Struktur[sunting | sunting sumber]
Gliserofosfolipid diperoleh daripada gliserol-3-fosfat dalam laluan de novo (dari kosong).[3] Istilah "gliserofosfolipid" menandakan sebarang terbitan asid gliserofosforik yang mengandungi sekurang - kurangnya satu sisa O-asil, atau O-alkil, atau O-alk-1'-enyl yang melekat pada bahagian gliserol. Kumpulan fosfat membentuk ikatan ester kepada gliserol. Hidrokarbon berantai panjang biasanya dilekatkan melalui kaitan ester dalam bakteria/eukariota dan oleh kaitan eter dalam archaea. Dalam bakteria dan prokariot, lipid terdiri daripada diester biasanya asid lemak C16 atau C18. Asid ini berantai lurus dan, terutamanya untuk ahli C18, boleh menjadi tidak tepu. Untuk archaea, rantai hidrokarbon mempunyai panjang rantai C10, C15, C20 dan lain-lain kerana ia berasal daripada unit isoprena. Rantai ini bercabang, dengan satu substituen metil setiap subunit C5. Rantai ini dikaitkan dengan gliserol fosfat melalui hubungan eter.[2] Dua rantai hidrokarbon yang melekat pada gliserol adalah hidrofobik manakala kepala kutub, yang terutamanya terdiri daripada kumpulan fosfat yang melekat pada karbon ketiga tulang belakang gliserol, adalah hidrofilik.[4] Ciri dwi ini membawa kepada sifat amphipathic glycerophospholipids.
Ia biasanya disusun menjadi dwilapisan dalam membran dengan kepala hidrofilik kutub melekat ke luar kepada persekitaran akueus, dan ekor hidrofobik bukan kutub menghala ke dalam.[5] Gliserofosfolipid terdiri daripada pelbagai spesies pelbagai yang biasanya berbeza sedikit dalam struktur. Struktur yang paling asas ialah fosfatida. Spesies ini merupakan perantaraan penting dalam sintesis banyak fosfogliserida. Kehadiran kumpulan tambahan yang melekat pada fosfat mewujudkan banyak fosfogliserida yang berbeza.
Mengikut tatanama, struktur sebatian ini menunjukkan 3 atom karbon gliserol secara menegak dengan fosfat melekat pada atom karbon nombor tiga (di bahagian bawah). Plasmalogen dan fosfatidat ialah contoh.[6]
Tatanama dan stereokimia[sunting | sunting sumber]
Secara umum, gliserofosfolipid menggunakan tatatanda "sn", bermaksud penomboran stereospesifik.[7] Apabila huruf "sn" muncul dalam tatanama, tatanama ini menunjukkan kumpulan hidroksil karbon kedua gliserol (2-sn) berada di sebelah kiri pada unjuran Fischer. Penomboran mengikut salah satu unjuran Fischer, iaitu 1-sn karbon di bahagian atas dan 3-sn di bahagian bawah.[8]
Kelebihan tatatanda khusus ini ialah konfigurasi ruangan (D atauL) molekul glisero ditentukan secara intuitif oleh bahagian di kedudukan sn-1 dan sn-3.
Contohnya, asid sn-glisero-3-fosforik dan sn-glisero-1-fosforik ialah enansiomer.
Kebanyakan minyak sayuran mempunyai asid lemak tak tepu dalam kedudukan 2, dengan asid lemak tepu dalam kedudukan 1 dan/atau 3.[7] Lemak haiwan lebih kerap mempunyai asid lemak tepu di 2, dengan asid lemak tak tepu dalam kedudukan 1 dan/atau 3.[7]
Contoh[sunting | sunting sumber]
Plasmalogen adalah sejenis fosfogliserida. Karbon pertama gliserol mempunyai rantai hidrokarbon yang disambungkan melalui hubungan eter, bukan ester. Sambungan lebih tahan terhadap serangan kimia daripada pautan ester. Atom karbon kedua (pusat) mempunyai asid lemak yang dihubungkan oleh ester. Karbon ketiga berpaut kepada etanolamin atau kolin melalui ester fosfat. Sebatian ini adalah komponen utama membran otot dan saraf.
Fosfatidat ialah sejenis lipid di mana dua atom karbon pertama gliserol ialah ester asid lemak, dan bahagian karbon ketiga adalah ester fosfat. Fosfat berfungsi sebagai penghubung kepada alkohol lain-biasanya etanolamina, kolina, serina atau karbohidrat. Identiti alkohol menentukan subkategori fosfatidat. Terdapat cas negatif pada fosfat dan, dalam kes kolina atau serina, ion ammonium kuaterner positif (serina juga mempunyai kumpulan karboksilat negatif). Kehadiran cas memberikan "kepala" dengan cas keseluruhan. Bahagian ester fosfat ("kepala") adalah hidrofilik, manakala baki molekul, asid lemak "ekor", adalah hidrofobik. Ini adalah komponen penting untuk pembentukan dwilapisan lipid.
Fosfatidiletanolamina, fosfatidilkolina dan fosfolipid lain ialah contoh fosfatidat.
Fosfatidilkolina ialah lesitin. Kolina ialah alkohol dengan ammonium kuaterner bercas positif yang terikat kepada fosfat dengan cas negatif. Lesitin terdapat dalam semua organisma hidup. Kuning telur mempunyai kepekatan tinggi lesitin yang penting secara komersial sebagai agen pengemulsi dalam produk seperti mayonis. Lesitin juga terdapat dalam otak dan tisu saraf.
Fosfatidilinositol membentuk komponen kecil sitosol dalam membran sel eukariotik dan memberikan molekul cas negatif. Kepentingannya bergantung pada peranannya dalam mengaktifkan reseptor deria yang berkait dengan fungsi rasa.
Fosfatidilserin adalah penting dalam pengisyaratan sel, khususnya apoptosis. Sel akan menggunakan fosfatidilserina ini untuk memasuki sel melalui mimikri apoptosis. Struktur lipid ini berbeza pada tumbuhan dan haiwan daripada aspek komposisi asid lemak. Di samping itu, fosfatidilserina memainkan peranan penting dalam kandungan otak manusia kerana ia membentuk 13-15% daripada fosfolipid dalam korteks serebrum manusia. Lipid ini terdapat di pelbagai tempat. Sebagai contoh, dalam diet manusia, kira-kira 130 mg berasal daripada fosfatidilserina. Ini telah dikatakan mempunyai kesan positif pada otak, kerana ia membantu mengurangkan tekanan dan memori yang lebih baik.[9]
Sfingomielin ialah sejenis sfingolipid yang mengandungi tulang belakang asas sfingoid. Ia boleh didapati dalam sarung mielin akson sel saraf dalam membran sel haiwan. Sfingomielin boleh didapati dalam telur atau otak lembu. Sfingolipid ini disintesis di retikulum endoplasma, dan diperkaya pada membran plasma dengan kepekatan yang lebih besar di lapisan luaran.[10]
Banyak fosfolipid lain yang wijid, sebahagian daripadanya ialah glikolipid. Glikolipid termasuk gula fosfatidil, di mana kumpulan berfungsi alkohol adalah sebahagian daripada karbohidrat. Gula fosfatidil terdapat dalam tumbuhan dan mikroorganisma tertentu. Karbohidrat sangat hidrofilik kerana banyaknya kumpulan hidroksil hadir.
Kegunaan[sunting | sunting sumber]
Fungsi dan kegunaan dalam membran[sunting | sunting sumber]
Glycerophospholipids adalah komponen struktur utama membran biologi. Sifat amphipathic mereka mendorong pembentukan struktur lipid dwilapisan membran. Membran sel yang dilihat di bawah mikroskop elektron terdiri daripada dua lapisan yang boleh dikenal pasti, atau "helaian", yang setiap satunya terdiri daripada deretan molekul gliserofosfolipid yang tersusun. Komposisi setiap lapisan boleh berbeza-beza bergantung pada jenis sel.
- Sebagai contoh, dalam eritrosit manusia, bahagian sitosol membran plasma terdiri terutamanya daripada fosfatidiletanolamina, fosfatidilserina dan fosfatidillinositol.
- Sebaliknya, bahagian eksoplasma (bahagian luar sel) terdiri terutamanya daripada fosfatidilkolina dan sfingomielin, sejenis sfingolipid.
Setiap molekul gliserofosfolipid terdiri daripada kumpulan kepala berkutub kecil dan dua rantai hidrofobik yang panjang. Dalam membran sel, dua lapisan fosfolipid disusun seperti berikut:
- ekor hidrofobik menghala satu sama lain dan membentuk lapisan tengah hidrofobik yang berlemak
- kumpulan kepala berion diletakkan di permukaan dalam dan luar membran sel
Selain daripada fungsinya dalam membran sel, ia berfungsi dalam proses selular lain seperti induksi dan pengangkutan isyarat. Berkenaan dengan isyarat, mereka menjadi pendahulu bagi prostaglandin dan leukotriena lain.[11] Pengedaran dan katabolisme khusus mereka membolehkan mereka menjalankan proses tindak balas biologi yang disenaraikan di atas.[12] Peranan mereka sebagai pusat penyimpanan pengutus kedua dalam membran juga merupakan faktor penyumbang kepada keupayaan mereka untuk bertindak sebagai pengangkut.[12] Ia juga mempengaruhi fungsi protein. Sebagai contoh, ia merupakan juzuk penting lipoprotein (protein larut yang mengangkut lemak dalam darah), justeru mempengaruhi metabolisme dan fungsinya.[5]
Pengemulsi[sunting | sunting sumber]
Gliserofosfolipid juga boleh bertindak sebagai agen pengemulsi untuk menggalakkan penyebaran satu bahan ke bahan lain. Ini kadangkala digunakan dalam pembuatan gula-gula dan pembuatan ais krim.
Dalam otak[sunting | sunting sumber]
Membran saraf mengandungi beberapa kelas gliserofosfolipid yang bertukar pada kadar yang berbeza berkenaan dengan struktur dan penyetempatan dalam sel dan membran yang berbeza. Terdapat tiga kelas utama iaitu; 1-alkil-2-asil gliserofosfolipid, 1,2-diasil gliserofosfolipid dan plasmalogen. Fungsi utama kelas gliserofosfolipid ini dalam membran saraf adalah untuk memberikan kestabilan, kebolehtelapan dan kecairan melalui perubahan khusus dalam komposisinya.[12] Komposisi gliserofosfolipid membran saraf sangat mengubah keberkesanan fungsinya. Panjang rantai asil gliserofosfolipid dan tahap ketepuan adalah penentu penting bagi banyak ciri membran termasuk pembentukan domain sisi yang kaya dengan asid lemak tak tepu. Penguraian gliserofosfolipid yang dimediasi reseptor oleh fosfolipase A(l), A(2), C, dan D menghasilkan penjanaan pengutus kedua seperti prostaglandin, eikosanoid, faktor pengaktif platelet dan diasilgliserol. Oleh itu, fosfolipid membran saraf menjadi takungan pengutus kedua. Mereka juga terlibat dalam apoptosis, modulasi aktiviti pengangkut dan enzim terikat membran. Perubahan ketara dalam komposisi gliserofosfolipid membran saraf telah dilaporkan berlaku dalam gangguan neurologi. Perubahan ini mengakibatkan perubahan dalam kecairan dan kebolehtelapan membran. Proses-proses, ini bersama dengan pengumpulan lipid peroksida dan metabolisme tenaga yang terjejas mungkin bertanggungjawab bagi kemerosotan saraf yang diperhatikan dalam gangguan neurologi.[13]
Metabolisme[sunting | sunting sumber]
Metabolisme gliserofosfolipid adalah berbeza dalam eukariot, sel tumor[14] dan prokariot. Sintesis dalam prokariot melibatkan sintesis asid fosfatidik gliserofosfolipid dan kumpulan kepala berkutub. Sintesis asid fosfatidik dalam eukariot adalah berbeza: terdapat dua laluan, satu kepada arah fosfatidilkolina (PC) dan fosfatidiletanolamina (PE). Gliserofosfolipid biasanya dimetabolismekan dalam beberapa langkah dengan perantaraan yang berbeza. Langkah pertama dalam metabolisme ini melibatkan penambahan atau pemindahan rantai asid lemak ke tulang belakang gliserol untuk membentuk perantaraan pertama, asid lisofosfatidik (LPA). LPA kemudiannya menjadi berasil untuk membentuk asid fosfatidik perantaraan (PA) seterusnya. PA boleh dinyahfosforilasi membawa kepada pembentukan diasilgliserol yang penting dalam sintesis PC.[5] PC adalah salah satu daripada banyak spesies gliserofosfolipid. Dalam laluan yang dipanggil laluan Kennedy, kepala berkutub ditambah untuk melengkapkan pembentukan keseluruhan struktur yang terdiri daripada kawasan kepala kutub, dua rantai asid lemak dan kumpulan fosfat yang melekat pada tulang belakang gliserol. Dalam laluan ini, kolina ditukar menjadi CDP-kolina yang mendorong pemindahan kumpulan kepala kutub untuk melengkapkan pembentukan PC. PC kemudiannya boleh ditukar kepada spesies lain seperti fosfatidilserina (PS) dan PE.[5]
Rujukan[sunting | sunting sumber]
- ^ Harayama, Takeshi; Riezman, Howard (May 2018). "Understanding the diversity of membrane lipid composition". Nature Reviews Molecular Cell Biology (dalam bahasa Inggeris). 19 (5): 281–296. doi:10.1038/nrm.2017.138. ISSN 1471-0080.
- ^ a b Caforio, Antonella; Driessen, Arnold J.M. (2017). "Archaeal phospholipids: Structural properties and biosynthesis" (PDF). Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1862 (11): 1325–1339. doi:10.1016/j.bbalip.2016.12.006. PMID 28007654.
- ^ Hishikawa, Daisuke; Hashidate, Tomomi; Shimizu, Takao; Shindou, Hideo (May 2014). "Diversity and function of membrane glycerophospholipids generated by the remodeling pathway in mammalian cells". Journal of Lipid Research. 55 (5): 799–807. doi:10.1194/jlr.R046094. ISSN 0022-2275. PMC 3995458. PMID 24646950.
- ^ Montealegre, Cristina; Verardo, Vito; Luisa Marina, María; Caboni, Maria Fiorenza (March 2014). "Analysis of glycerophosphate- and sphingolipids by CE". Electrophoresis. 35 (6): 779–792. doi:10.1002/elps.201300534. PMID 24301713.
- ^ a b c d Ecker, Josef; Liebisch, Gerhard (April 2014). "Application of stable isotopes to investigate the metabolism of fatty acids, glycerophospholipid and sphingolipid species". Progress in Lipid Research. 54: 14–31. doi:10.1016/j.plipres.2014.01.002. PMID 24462586.
- ^ Yong-Mei Zhang and Charles O. Rock (2008). "Thematic Review Series: Glycerolipids. Acyltransferases in bacterial glycerophospholipid synthesis". J Lipid Res. 49 (9): 1867–1874. doi:10.1194/jlr.R800005-JLR200. PMC 2515527. PMID 18369234.
- ^ a b c "Effects of Plant Oil Interesterified Triacylglycerols on Lipemia and Human Health". International Journal of Molecular Sciences. 19 (1): E104. 2017. doi:10.3390/ijms19010104. PMC 5796054. PMID 29301208.
- ^ Moss G.P. (www version) (1976). "Nomenclature of Lipids, Recommendations 1976". IUPAC-IUB Commission on Biochemical Nomenclature (CBN). Part I. Fatty Acid, Neutral Fats, Long-Chain Alcohols and Long-Chain Bases § Lip-1.13. Dicapai pada 27 Sep 2023.
- ^ "Scientific Opinion on the substantiation of health claims related to phosphatidyl serine (ID 552, 711, 734, 1632, 1927) pursuant to Article 13(1) of Regulation (EC) No 1924/2006 | EFSA". www.efsa.europa.eu (dalam bahasa Inggeris). 2010-10-19. Dicapai pada 2023-11-29.
- ^ Testi, Roberto (1996-12-01). "Sphingomyelin breakdown and cell fate". Trends in Biochemical Sciences. 21 (12): 468–471. doi:10.1016/S0968-0004(96)10056-6. ISSN 0968-0004.
- ^ Hermansson, Martin; Hokynar, Kati; Somerharju, Pentti (July 2011). "Mechanisms of glycerophospholipid homeostasis in mammalian cells". Progress in Lipid Research. 50 (3): 240–257. doi:10.1016/j.plipres.2011.02.004. PMID 21382416.
- ^ a b c Farooqui, AA; Horrocks, LA; Farooqui, T (June 2000). "Glycerophospholipids in brain: their metabolism, incorporation into membranes, functions, and involvement in neurological disorders". Chemistry and Physics of Lipids. 106 (1): 1–29. doi:10.1016/s0009-3084(00)00128-6. PMID 10878232.
- ^ Garcia, Christina (2011-06-30). "Metabolism of glycerophospholipids". We Sapiens.org. Diarkibkan daripada yang asal pada 2012-03-23.
- ^ "Glycerophospholipid synthesis as a novel drug target against cancer". Current Molecular Pharmacology. 4 (3): 167–175. November 2011. doi:10.2174/1874467211104030167. PMID 21222647.
Pautan luar[sunting | sunting sumber]
- Glycerophospholipids dalam Tajuk Subjek Perubatan (MeSH) di Perpustakaan Perubatan Negara AS
- Diagram at uca.edu
{kind=link}